
Copa do mundo, comportamento animal e o triptofano
Artigo de Lívea Maria Gomes – Departamento de Nutrição da König do Brasil, publicado na Revista Vet Science
Como todo bom brasileiro, sabemos que estamos a praticamente um mês da Copa do Mundo FIFA 2018 e junto com ela e toda expectativa sobre a nossa seleção, teremos muitas comemorações (ou não), gritos de gol, buzinas, cornetas, som alto e fogos de artifício!
Essa data também nos abre espaço para reflexão e diálogo sobre Bioética considerando os interesses de todas as partes, o papel da sociedade e os efeitos destas atividades de diversão e lazer que ocasionam, de forma quase que inconsciente, inúmeros problemas ambientais e de bem-estar aos animais.
Os fogos de artifício são utilizados para diversas finalidades em situações comemorativas e simbólicas desde o século XIV, quando os árabes os levaram para Europa. Apesar de seu som e brilho serem muito apreciados pelos humanos são fonte de perturbação para inúmeras espécies de animais silvestres e domésticas, dentre os quais estão os cães e gatos que tem muita sensibilidade auditiva.O Brasil é o segundo maior produtor de fogos de artifício, perdendo apenas para a China. (CAPILÉ et al., 2014).
O animal, quando exposto a estes sons e movido pelo medo, procura se afastar do estímulo estressor, tentando se esconder dentro ou embaixo de móveis ou espaços restritos; busca pessoas para se proteger; pode tentar fugir pela janela; cavar buracos; ficar agressivo; apresentar desorientação, salivação excessiva e respiração ofegante; tremedeira; diarreia temporária; e urinar ou defecar involuntariamente. Há também possibilidade de ocorrer acidentes durante a tentativa de fuga, tais como atropelamentos, quedas, colisões ou o desaparecimento do animal, que pode percorrer longas distâncias em estado de pânico e depois não conseguir retornar ao seu local de origem (BLACKWELL et al., 2013).
Esses estímulos sonoros são reconhecidos pelo sistema auditivo e avaliados quanto a sua relevância, para que o indivíduo possa reconhecer situações de perigo e então desencadeiam respostas fisiológicas de estresse agudo por meio da ativação do sistema neuroendócrino, que eleva rapidamente os níveis de cortisol rapidamente em busca de adaptação imediata após a exposição ao estímulo estressor e o retorno aos níveis basais em uma hora ou mais (WENDELAAR BONGA, 1997; BEERDA et al., 1999).
Os problemas recorrentes aos danos causados pelo medo dos fogos de artifício podem variar desde estresse e ansiedade crônicos, insuficiência cardíaca pós convulsão, crises de bronquite aguda, abortos, lesões fatais e necessidade do uso de sedativos pelos animais.
Segundo Beaver (2001), 20% dos cães com fobia de ruídos possuem problemas suficientemente graves a ponto de seus proprietários procurarem ajuda profissional. Blackwell e colaboradores (2013) relataram que entre os 383 proprietários entrevistados sobre reações de medo a ruídos apresentados pelos seus cães, 29% dos proprietários procuraram ajuda profissional.
Apesar do tratamento majoritariamente adotado pelos veterinários ser o uso de sedativos, existem opções de nutrientes que podem antecipadamente serem adicionados a dieta do animal com a intenção de modelar a resposta ao estresse a qual os animais serão expostos. Neste sentido, foram desenvolvidos estudos com o objetivo de avaliar os efeitos dos nutrientes das dietas sobre a modulação do comportamento em cães e gatos. Dentre os nutrientes que apresentam maior importância e estudos mais consistentes, destaca-se o aminoácido triptofano.
O triptofano, um aminoácido essencial para cães e gatos, é precursor de compostos bioativos, limitante para síntese de proteínas e deve ser fornecido por meio da alimentação, visto que sua síntese não pode ser realizada pelo animal (PEREIRA, 2007). Quando o triptofano é suplementado na dieta além da exigência para síntese proteica, pode ser utilizado como suplemento terapêutico (LI et al., 2006).
O triptofano é facilmente absorvido pelos capilares da parede intestinal, no qual a maior parte dele – cerca de 80-90%, é transportado no sangue pela albumina, diferentemente de outros aminoácidos (PARDRIDGE, 1979), assim, apenas 10-20% de triptofano está presente como forma livre no plasma. O transporte do triptofano pode ser alterado pela concorrência exercida por outros aminoácidos, como valina, leucina, isoleucina, fenilalanina, tirosina e metionina, que se ligam aos mesmos transportadores (HENRY et al., 1992; BOISEN et al., 2000; PEREIRA, 2007). Portanto, a eficiência da utilização de determinado aminoácido é dependente da concentração adequada de outros aminoácidos e do teor proteico da dieta (HENRY et al., 1992; MILLET, 2012). Em contrapartida, a dieta rica em carboidratos proporcionará aumento de triptofano utilizado pelo cérebro (LE FLOC’H e SÈVE, 2007). Para maior produção de serotonina pelo cérebro é necessária uma quantidade maior tanto de triptofano em forma livre quanto de triptofano ligado a proteínas da dieta. O L-triptofano presente em suplementos alimentares eleva rapidamente o seu nível plasmático, enquanto que o triptofano ligado a proteínas da dieta leva mais tempo para aparecer na corrente sanguínea e afetar a produção de serotonina (SÈVE, 1999).
Os compostos provenientes do triptofano são formados em duas rotas ou vias metabólitas (Figura 1): via serotoninérgica e via das quinureninas, essas rotas são dependentes de quantidades adequadas de biopterina, magnésio e piridoxina (B6) para que haja a conversão deste aminoácido em seus subprodutos (PAREDES et al., 2009).
A via serotoninérgica, ativada principalmente em plaquetas, no intestino e neurônios de nervos serotoninérgicos, converte o triptofano em serotonina em duas etapas: a conversão do triptofano em 5-hidroxitriptofano pela enzima triptofanohidroxilase (TPH) e a descarboxilação do 5-hidroxitriptofano em serotonina (5-HT ou 5-hidroxitriptamina) (LE FLOC’H et al., 2011). A contribuição das vias metabólicas do triptofano pode ser diferente de acordo com o estado fisiológico e patológico, além disso, a proporção de triptofano a ser metabolizado a serotonina ou quinurenina é totalmente dependente da capacidade de síntese proteica (LE FLOC’H et 23 al., 2011).
Em média de 4 a 6% do triptofano da dieta sofre degradação bacteriana no intestino, 0,5% é excretado na urina e apenas 1% a 2% é convertido na via serotoninérgica (BENDER, 1983). Wolf (1974) afirmou que menos de 1% do triptofano ingerido será convertido em serotonina. A enzima triptofanohidroxilase que limita a taxa de síntese de serotonina a partir do triptofano, tem atividade parcial de sua capacidade, de tal modo que o aumento do triptofano disponível pode dobrar a taxa de síntese de serotonina (CARLSON, 1977; HEINE et al., 1995; SAINIO et al., 1996). O transporte e a disposição do triptofano parecem ser fatores limitantes para a síntese de serotonina pelo cérebro (PARDRIDGE, 1998; LE FLOC’H e SÈVE, 2007).
TRIPTOFANO E ESTRESSE
O mecanismo pelo qual o triptofano pode interagir com a resposta ao estresse não é totalmente elucidado, porém sabe-se que está envolvido com a produção e turnover de serotonina no cérebro (LEATHWOOD, 1987). A serotonina ou 5-hidroxitriptamina (5-26 HT) é uma monoamina biogênica de baixo peso molecular pertencente ao grupo indol que funciona como neurotransmissor inibitório do sistema nervoso central (PEREIRA, 2007). Desenvolve papel fundamental na regulação de processos comportamentais e fisiológicos dos animais, tais como regulação de temperatura, apetite (ZHANG et al., 2007), regulação das funções gastrintestinais, hemodinâmica (LE FLOC’H et al., 2011), secreção de hormônios do estresse (ADEOLA et al., 1993; LEPAGE et al., 2003; KOOPMANS et al., 2005), imunidade (MELCHIOR et al., 2004; LE FLOC’H et al., 2009), regulação do ritmo circadiano, comportamento agressivo (CORTAMIRA et al., 1991) e sexual, sensibilidade à dor, humor (MARKUS et al., 2000) e bem-estar (MEUNIER-SALAÜN et al., 1991; SHEN et al., 2012).
A serotonina não é capaz de atravessar a barreira sanguínea do cérebro, portanto, os seus efeitos dentro do sistema nervoso central (SNC) dependem da transferência de triptofano por entre essa barreira. Uma vez dentro do SNC, o triptofano é facilmente convertido a serotonina (LI et al., 2006) e após sua síntese, a serotonina é liberada a partir das vesículas sinápticas para a fenda sináptica, onde se liga aos seus receptores específicos. A deficiência de triptofano prejudica a síntese da serotonina no cérebro (HENRY et al., 1996). O sistema serotonergico reage a fatores estressantes (MASON, 1968; LEPAGE et al., 2005) e, frente ao ao estresse há maior turnover da serotonina pelo cérebro (CHAULOFF, 17 1993; BAGDY, 1995) possibilitando melhor resposta adaptativa (DEAKIN e GRAEFF, 1991; GRAEFF et al., 1996). A liberação da serotonina durante o período em que o animal se encontra em vigília atua também como modulador positivo na produção do hormônio melatonina (PAREDES et al., 2009), pois, alguns tecidos são capazes de utilizá-la para essa síntese numa via intermediada por N-acetilserotonina.
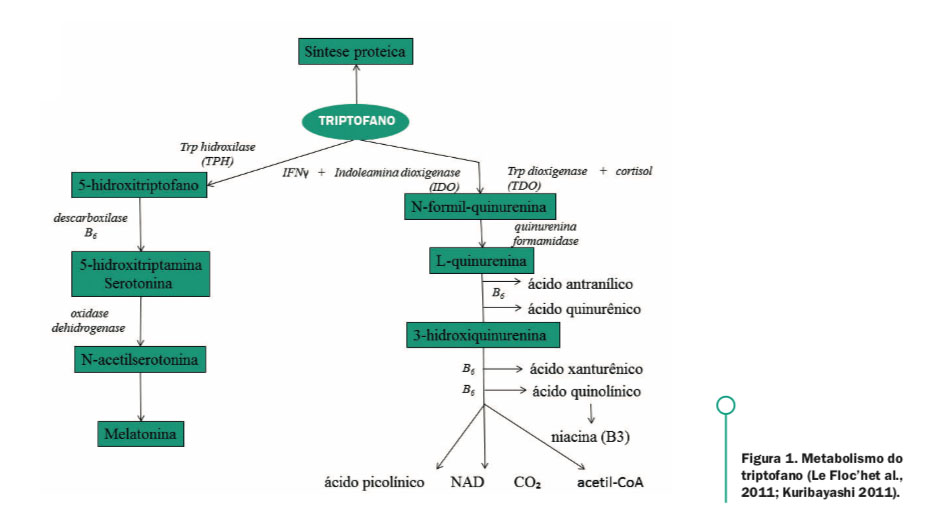
Figura 1. Metabolismo do triptofano (Le Floc’het al., 2011; Kuribayashi 2011).
Em teoria, o aumento das concentrações dietéticas de triptofano pode ser usado como estratégia nutricional e farmacológica para controlar o estresse e comportamentos agressivos elevando os níveis circulantes de serotonina e melatonina no organismo (ADEOLA e BAL, 1992; VAN HIERDEN et al., 2004). O uso do triptofano na dieta pode reduzir a agressividade e aliviar o estresse em muitas espécies, tais como humanos (MARKUS et al., 1998; ATTENBURROW et al., 2003), suínos (MEUNIER-SALAÜN et al., 1991; ADEOLA e BAL, 1992; PEREIRA et al., 2008; SÈVE., 1999; POLETTO, 2010), roedores (GIBBONS et al., 1979; LEATHWOOD, 1987 ), aves (SHEA et al., 1990; LAYCOCK e BALL, 1990; SHEA-MOORE et al., 1996; ROSEBROUGH, 1996;SAVORY et al., 1999; VAN HIERDEN et al., 2004) e peixes (WINBERG et al., 2001; LEPAGE et al., 2002). Além disso, para cavalos, altas dosagens de triptofano foram propostas para efeito sedativo (GRIMMETT e SILLENCE, 2005).
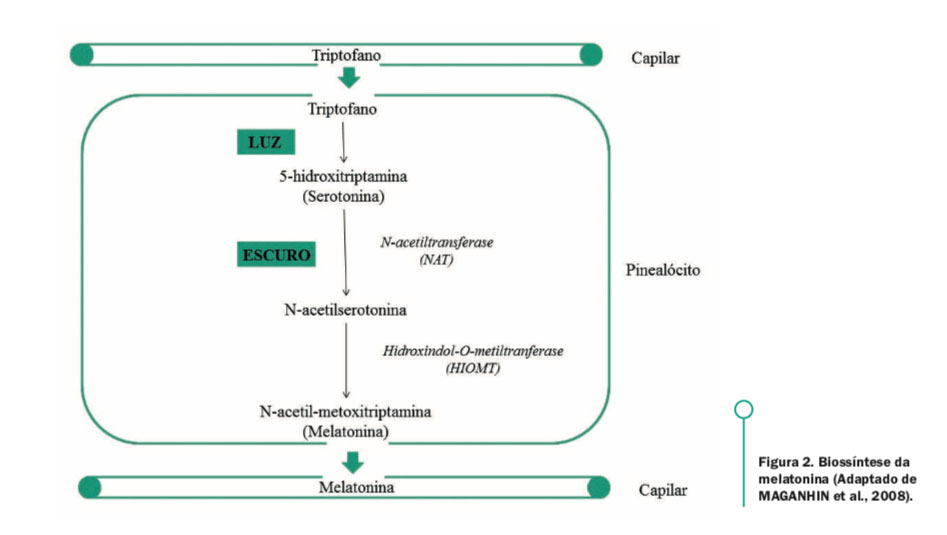
Figura 2. Biossíntese da
melatonina (Adaptado de
MAGANHIN et al., 2008).
Para que a suplementação de triptofano seja modeladora da resposta ao estresse e da sensibilidade dos animais, são necessárias altas doses farmacológicas ou a deficiência deste aminoácido (KOOPMANS et al., 2005; GUZIK et al., 2006), visto que sua suplementação na dieta em doses que apenas atendam as exigências nutricionais ocasiona efeitos mais discretos ou inexistentes (MEUNIER-SALAÜN et al., 1991).
Em suínos, Koopmans et al. (2005) e Guzik et al. (2006) observaram que a suplementação de triptofano na dieta é capaz de diminuir as concentrações de cortisol e noradrenalina nos animais antes do estresse e menores concentrações de cortisol, noradrenalina e adrenalina após a exposição dos mesmos ao estresse, indicando que o triptofano pode reduzir respostas ao estresse.
Em cães e gatos, os estudos que avaliem a suplementação de triptofano e sua relação com as variáveis comportamentais são relativamente recentes.
Na avaliação de atividade elétrica cerebral de cães, Elmor (2009) observou redução nas bandas alfa, beta, delta e teta, de cães com peso médio de 32,5kg, suplementados com 1,11g/dia de triptofano, a porcentagem de triptofano em relação aos demais aminoácidos (fenilalanina, tirosina, leucina, isoleucina, valina) foi 7,5 vezes maior na dieta suplementada em comparação à dieta não suplementada, levando a supor que a suplementação com o aminoácido modula a atividade elétrica cerebral dos animais, tendo positivo efeito sobre a redução do estresse dos animais.
Dessa forma, pode-se concluir que a suplementação com triptofano pode ser uma opção para melhorar o bem-estar de cães e gatos expostos a eventos estressantes, não só durante a Copa do Mundo, mas também em datas comemorativas distribuídas ao longo do ano como partidas de futebol, celebrações religiosas, Natal, Ano Novo e Festas Juninas.
LÍVEA MARIA GOMES – Zootecnista
Formada pela FZEA-USP (Pirassununga/SP), pós-graduada em Controle de Qualidade na Indústria de Alimentos pela Universidade de Sorocaba (Sorocaba/ SP) e mestre em Nutrição e Produção Animal pela FMVZ-UNESP (Botucatu/SP) – Departamento de Nutrição da König do Brasil LTDA
Acesse o artigo original no site da Revista Vet Science.
CONFIRA AS REFERÊNCIAS BIBLIOGRÁFICAS NO CONTEÚDO ONLINE COMPLEMENTAR EM WWW.REVISTAVETSCIENCE.COM.BR






Deixe uma resposta
Want to join the discussion?Feel free to contribute!